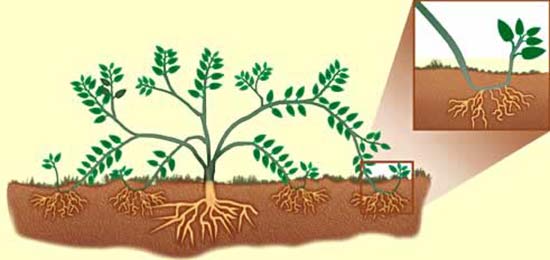
Опыление
Содержание:
- Самоопыление. искусственное опыление.
- Селекция и самоопыление
- Описание механизма
- Литература
- Формы перекрестного опыления
- Примеры в окружающей природе
- Механизмы стимулирования перекрестного опыления
- О продолжительности сазаньей тусовки
- Способы привлечения опылителей
- Шаг второй. Изучение исходного материала
- Виды перекрестного опыления
- Приспособления по недопущению самоопыления[править | править код]
- Особенности анемофильных растений
- Генеративный орган высших растений
Самоопыление. искусственное опыление.
Кроме перекрестного опыления, у некоторых растений происходит самоопыление. При самоопылении пыльца попадает с тычинок на рыльце пестика того же цветка. Перечислим некоторые растения-самоопылители: пшеница, лен, ячмень, горох, фасоль, картофель и некоторые другие. Самоопыление у растений чаще всего происходит в закрытом цветке. Когда цветок раскрывается, пыльца уже оказывается прилипшей к рыльцу пестика.
Перекрестное опыление насекомыми и ветром, а также самоопыление растений — это естественное опыление. Известно и искусственное опыление.
При искусственном опылении человек с определенной целью сам переносит пыльцу с тычинок на пестики цветков. Например, чтобы получить новый сорт, ученые-садоводы искусственно переносят пыльцу с тычинок одного сорта растений на пестики цветков другого сорта.
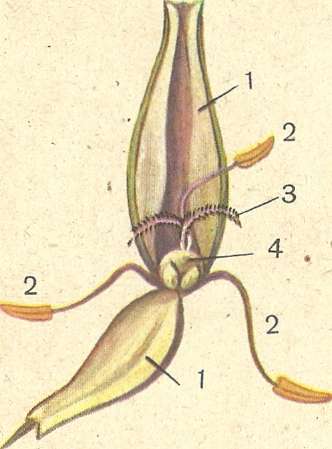
Рис. 96. Строение цветка ржи : 1 — цветочная чешуя; 2 — тычинки; 3— рыльце пестика; 4 — завязь.

Рис. 98. Сбор пыльцы для искусственного опыления кукурузы.
Перенести пыльцу можно чистой сухой акварельной кисточкой или кусочками резинки, прикрепленной к проволочке. Цветки, на которые будет перенесена пыльца, надо заранее подготовить к опылению. Для этого из них еще в бутонах удаляют тычинки, чтобы не произошло самоопыления. Затем закрывают цветки марлевым мешочком, чтобы ветер или насекомые случайно не занесли пыльцу на пестик цветка. Когда распустятся бутоны, помещенные в марлевые мешочки, на пестики цветков переносят заблаговременно заготовленную пыльцу.
Искусственное опыление иногда применяют для повышения урожайности культурных растений. Примером может служить дополнительное искусственное опыление ржи. Его проводят в тихую, безветренную погоду, потому что без ветра пыльца не разносится. Делается это так. Два человека идут по полю с натянутой веревкой, наклоняя ею цветущие растения. При этом пыльца высыпается и попадает на рыльца пестиков. В результате дополнительного опыления урожай ржи повышается.
При искусственном опылении повышается урожай подсолнечника и кукурузы. Подсолнечник опыляют в холодную погоду. Дело в том, что подсолнечник — перекрестноопыляемое растение. Его опыляют насекомые, но в плохую погоду их летает мало. Чтобы опыление произошло и
без насекомых, человек проходит вдоль каждого ряда растений, прижимает варежку из мягкой материи к корзинке цветков одного подсолнечника. Затем переносит варежкой пыльцу на соцветия другого.
А как искусственно опылить кукурузу? Это перекрестноопыляемое растение. Тычиночные цветки кукурузы собраны в соцветие, называемое метелкой. Пестичные цветки образуют початки , растущие в пазухах листьев на средней части стебля. Во время цветения из початка свисают длинные нитевидные столбики с рыльцами. Ветер переносит на них пыльцу из тычиночных соцветий. Тычиночные цветки созревают раньше, чем пестичные. Часто в безветренную погоду пыльца осыпается, не попадая на рыльца. Многие пестичные цветки отцветают, не опылившись. Урожай зерна кукурузы снижается.
Чтобы искусственно опылить кукурузу, собирают пыльцу, стряхивая ее с метелки с созревшей пыльцой в воронку из плотной бумаги. Затем собранную пыльцу наносят на рыльца пестичных цветков.
Початки, завязавшиеся в результате искусственного опыления, бывают более крупными. Семена в них особенно хороши.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Селекция и самоопыление
Это явление широко используется в селекции растений. Нам известно, что самооплодотворение и скрещивание близкородственных организмов ведет к переходу генов в гомозиготное состояние и приводит к снижению жизнестойкости и урожайности, а впоследствии и к вырождению. Непрерывный процесс мутаций, которые накапливаются, большинство из которых рецессивны и неблагоприятны, – причина этого угнетения. У растений с перекрестным опылением эти мутации находятся в гетерозиготном состоянии и никак не проявляются. При самоопылении вероятность их перехода в гомозиготу во много раз возрастает, но они не сохраняются в популяции из-за естественного устранения. Самоопыление в селекции используется как инструмент создания чистых (гомозиготных) линий с закрепленными признаками. Несмотря на снижение продуктивности, после гибридизации часто появляется явление гетерозиса – силы гибридов от сортов с самоопылением. Это явление называется межлинейной гибридизацией, и в магазинах мы можем увидеть именно такие гибридные семена (они помечены символом F1). В первом поколении гибриды превосходят чистые линии по урожайности, но в дальнейших поколениях эффект силы гибридов исчезает.
Описание механизма
Цветковые растения получили своё название, потому что имеют морфологический орган — цветок, способный к половому размножению и привлечению опылителей. Для образования завязей, формирования плодов и развития семян первым условием является опыление, то есть перенос частиц пыльцы. Природа позаботилась о том, чтобы всё проходило успешно.
Это может осуществляться ветром — тогда целые облака невесомых пылинок поднимаются и передвигаются в потоках зефира. Многие деревья (дубы, ясени и сосны), сельскохозяйственные злаковые и кукуруза «заботятся» об увеличении вероятности попадания на нужные рыльца, вырабатывая заведомо большие количества пыльцы. Её частички очень лёгкие, почти невесомые, чтобы беспрепятственно «проплывать» по воздуху, а у некоторых имеются газовые пузырьки, способствующие более долгому путешествию.
Но чаще всего опыление осуществляют разные насекомые
Чтобы привлечь внимание, растения окрашиваются в очень яркие цвета и источают сильный аромат. Если их размеры малы, они группируются в пышные соцветия или окружают себя разноцветными листьями — прицветником, как это делает мексиканская красавица пуансеттия
По сравнению с теми, что опыляются ветром, пыльцевые зёрна таких цветов обычно более крупные, шероховатые и клейкие, чтобы уверенно прилипать к насекомым.
Для привлечения опылителей медовые железы растений специально выделяют нектар — сок, богатый различными сахарами (в основном это сахароза и фруктоза). Кроме того, в состав входят:
- кислоты (аспарагиновая и глютаминовая);
- минеральные соли;
- ферменты;
- ароматические компоненты.
Осы, бабочки и шмели охотно употребляют нектар в пищу. Медоносные пчёлы, собирая и пряча его в соты улья, производят мёд. Акации специально выделяют сладкий секрет для привлечения муравьёв, защищающих древесину от зубов травоядных животных. Спектр опылителей может быть широким (эуфилия), подчиняться опылению несколькими родственными или только определённой жизненной формой (олигофилия) или требовать один вид насекомых (монофилия).
Если говорят, что опылитель обладает полилектией, это свидетельствует о чрезвычайно высоком уровне приспособленности к опылению, он способен обслуживать представителей различных семейств. Посещение ограниченной группы, состоящей из одного семейства или растений с однотипными соцветиями, свидетельствует о наличии олиголектии. В случае монолектии опылитель обязательно питается одним видом или родом растений и опыляет только их.
После этого наступает очередь оплодотворения, которое происходит у всех по-разному: чаще период длится несколько недель, но иногда для полноценного слияния половых клеток требуются месяцы. Пыльца, находящаяся на рыльце, для этого должна созреть, обладать достаточной жизнестойкостью и иметь сформированный женский гаметофит (зародышевый мешок). В него и проникает пыльцевая трубка, растущая в направлении завязи через семенной зачаток.
Оказавшись возле яйцеклетки, она спешит разорваться и высвободить два спермия: один соединяется с яйцеклеткой и даёт жизнь зародышу, второй сливается с диплоидным ядром, образуя триплоидную клетку и формируя эндосперм. Такое двойное оплодотворение позволяет получить семя, защищённое кожурой, а из завязи затем возникнет желанный плод.
Литература
- Пономарёв А. Н., Демьянова Е. И., Грушвицкий И. В. Опыление // Жизнь растений. В 6-ти т. / под ред. А. Л. Тахтаджяна. — М.: Просвещение, 1980. — Т. 5. Ч. 1. Цветковые растения. — С. 55—78. — 430 с. — 300 000 экз.
- Суриков И. М. Несовместимость и эмбриональная стерильность растений. — М.: ВАСХНИЛ, Агропромиздат, 1991. — 220 с.
- Коровкин О. А. Анатомия и морфология высших растений: словарь терминов. — М.: Дрофа, 2007. — С. 35, 52, 89, 137, 162. — 268, с. — (Биологические науки: Словари терминов). — 3000 экз. — ISBN 978-5-358-01214-1.
- Современный словарь иностранных слов: Ок. 20000 слов. — 4-е изд., стер. — М.: Русский язык, 2001. — С. 139, 150, 209, 323, 496, 497. — 742 с. — Доп. тираж 10 000 экз. — ISBN 5-200-02989-9.
- Демьянова Е. И. Антэкология: учеб. пособие по спецкурсу / Перм.гос. ун-т. — Пермь, 2010. — 116 с.: ил. — ISBN 978-57944-1581-0
Формы перекрестного опыления
В ходе эволюционной истории формы передачи мужских гамет развивались в этом порядке, при этом водные растения ( водоросли ), из которых развились сосудистые растения, были не пыльцой, а плавающими гаметами. В мха , в котором гаметы , передаваемые по воде, не пыльцы либо. Только семенные растения вырабатывали пыльцу в форме переноса пыльцы .
- Абиотическое опыление
- Биотическое опыление
Водоросли растут под водой
Количество пыльцы Dactylis glomerata
Pigeon хвост вставляет свой ствол в трубчатый цветок в георгин .
Колибри с фиолетовыми ушами
Летать на муховике
-
- ( Вода ) гидрофильность
- ( Ветер ) анемофилия
- ( Животные ) зоофилия
- (Человеческая) антропофилия
Опыление воды
Редкое опыление воды происходит у некоторых растений, которые растут под водой или на поверхности воды. Пыльца может переноситься над или под поверхностью воды.
Опыление ветром
При ветровом опылении пыльца передается ветром и случайно попадает на рыльце другого цветка.
У опыляемых ветром растений часто бывает незаметный цветочный покров или он полностью отсутствует. Нектар и отдушки не производятся. Цветки часто располагаются в многоцветковые, часто однополые соцветия . Обильное количество пыльцы часто образуется на длинных тычинках, движущихся на ветру. Цемент пыльцы обычно отсутствует. Шрамы большие и сильно разделены.
Типичными ветровыми опылителями являются травы , пшеница , рожь и кукуруза . Пыльца ветроопыляющих растений может вызвать сенную лихорадку .
Опыление животными
Животные опыление подразделяются в зависимости от типа опыления животных: наиболее часто встречающихся животными опыления насекомыми. Когда опыление насекомыми можно дифференцировать с помощью Fliegenblütigkeit , Bienenblütigkeit , Tagfalterblütigkeit и других.
В тропиках важно опыление птицами, равно как и опыление летучими мышами. Цветки, опыляемые птицами, так называемые птичьи цветы , часто имеют ярко-красный цвет, одноцветные
насекомые не видят.
Способы, которыми растения привлекают своих опылителей, разнообразны. Многие из опыляемых насекомыми растений опыляются насекомыми, пьющими нектар и / или собирающими пыльцу, такими как пчелы , шмели , бабочки или журчалки . Животных обычно привлекает большой и ярко окрашенный цветочный покров . Часто цветок оформлен дорсивентрально . Если присутствуют нектар и ароматы, говорят о цветках нектара . Пока животные собирают нектар, их присыпают пыльцой. Если они летят к следующему цветку, пыльца прилипнет к их рыльцу. Цветущие растения без нектара и отдушек называют цветками пыльцы . При адаптации к опылению насекомыми тычинки часто короче, а рыльце не очень разделено.
Относительно основных опылителей у растений развились определенные приспособительные характеристики:
Нектар растений, опыляемых бабочками , часто находится на дне длинных трубок, в которые бабочки вставляют свой хоботок. Если моль — главные опылители, цветы часто раскрываются только вечером. Такие цветы обычно не бросаются в глаза, но обладают интенсивным ароматом.
У растений, опыляемых преимущественно мухами, преобладают мелкие нектарники . Здесь характерен характерный грибной или падалный запах .
В частности , орхидеи разработали специальные механизмы для привлечения насекомых-опылителей. Некоторые виды не предлагают нектар, а скорее имитируют цветы других растений, которые дают нектар через форму и цвет прицветников. Некоторые виды орхидей привлекают самцов определенных видов насекомых феромонами и побуждают их к совокуплению (например, виды амброзии ) → обманчивый цветок .
У цветов-ловушек для чайников есть, например, Aristolochia , Yellow Lady’s Slipper и Arum . Из-за особой структуры цветка мелкие насекомые попадают в чашеобразное расширение цветка или, в случае арума, на покров и могут покинуть его только через такие устройства, как ловушка для волосков, когда произошло опыление.
Искусственное опыление (человек)
Чтобы предотвратить нежелательное случайное опыление при селекции растений , существуют методы искусственного опыления. Для этого пыльцу выбранного родительского растения можно перенести на рыльце цветка выбранного родительского растения, например, с помощью тонкой кисти . Грегор Мендель уже использовал этот метод в своих экспериментах по разведению цветковых растений. Мендель удалил тычинки на ранней стадии, чтобы исключить нежелательное самоопыление или случайное перекрестное опыление, и он мог быть уверен, что потомство происходит от выбранных родительских растений.
Примеры в окружающей природе
От опыления зависит сохранение вида, поэтому так важно, чтобы всё проходило успешно. Растения, в отличие от животных, не могут самостоятельно передвигаться в поисках партнёра для размножения и вынуждены уповать на помощь союзников — ветра, воды или живых существ
Самоопыляющимся растениям присущи следующие отличительные признаки:
- цветки не имеют запаха и не выделяют нектара;
- пестики располагаются ниже тычинок;
- пыльца созревает ещё на стадии бутонизации, и опыление осуществляется в нераспустившемся цветке, как у гузмании или арахиса.
Томаты «умеют» самоопыляться факультативно: их тычинки оказались сросшимися, поэтому пестик без труда оплодотворяется своей пыльцой. Малопрогрессивному потомству, полученному таким образом, угрожает вырождение. Этого не случится, если некоторые цветки подвергнутся внутривидовому опылению и приобретут несколько иные родительские зачатки. Тогда они будут обладать более высокими способностями к приспособлению и выживут в процессе естественного отбора.
Мужские деревья двудомных тополя и облепихи интересны только цветками с пыльцой, а женские особи плодоносят. В случае тополя пушистые семена — продукт жизнедеятельности женских деревьев. Если высаживать для озеленения исключительно мужские черенки, можно навсегда избавиться от надоедливого пуха. Оранжевые плоды облепихи дают только женские кусты, но если рядом не посадить мужское растение, то урожая не будет, а обычное соотношение составит 10:1.
Однополые цветки однодомной кукурузы собраны в разнотипные соцветия: мужские — «метёлка» на макушках, женские — «початок» в листовых пазухах. Другие характерные представители однодомных — тыква и огурец — тоже образуют цветки разного типа в пределах одного растения. По внешним признакам они почти неотличимы, но после опыления мужские сразу отмирают и осыпаются, а из оплодотворённых женских будут формироваться завязи.
Механизмы стимулирования перекрестного опыления
Карл Фрич все еще полагал, что в животном мире не существует самооплодотворения . Ему было известно самоопыление растений, и он обладает морфологическими характеристиками и особенностями во временной последовательности, описанной Blühvorgangs, которые снижают вероятность самоопыления и способствуют перекрестному опылению. В учебнике ботаники из Страсбургер и других учебниках эти механизмы описаны подробно. Наиболее распространены:
Формирование однополых цветков
В случае ветровых опылителей они бывают одно- или двухкомпонентными . Пространственно разделенные однополые цветки ( диклина ) также поддерживают перекрестное опыление. У моноэа на одном экземпляре растения присутствуют женские и мужские цветки. В случае раздельнополых цветов бывают мужские и женские особи. Следовательно, у одиночной особи есть только мужские или женские цветки. К этой группе растений относятся облепиха , ива обыкновенная или капуста обыкновенная . Поскольку мужские и женские цветки распределены между разными особями, самоопыление невозможно.
Герогамия
Магнолия с пространственно разделенными гинецеем и андроцеем
Если тычинки и рыльца внутри цветка несколько более пространственно разделены морфологическими особенностями, это называется геркогамией . Примером этого является радужная оболочка глаза . У многих из этих цветов клеймо поднимается гинофором таким образом, что несколько снижается вероятность того, что на рыльце попадут пыльцевые зерна с собственного цветка.
Другой стилус
У некоторых видов растений есть особи, у которых стили длинные, а пыльники глубоко посажены, и особи, у которых стили короткие, а пыльники высоко посажены. Итак, есть два разных типа цветов. Это называется разным стилусом или гетеростилией . У других видов с двумя тычинковыми кругами бывает даже три разных типа цветков, в зависимости от того, находится ли стиль на нижнем, среднем или верхнем уровне ( тристилий ).
Оплодотворение происходит только тогда, когда пыльца переходит с одного уровня на клеймо на том же уровне. Например, пыльца с коротких тычинок также должна попасть на рыльце короткого стилуса. Но этого не может произойти в цветке.
Примерами являются примула с двумя уровнями или семейство вербейников пурпурных и семейства щавелевых, в которых встречается тристилий.
Пре-мужественность
Есть также растения, препятствующие самоопылению с временным отрывом. Когда Vormännlichkeit ( Proterandrie ) пустые пыльники собирают пыльцу до того, как рыльце того же цветка готово к зачатию. Это происходит, например, с семейством маргариток , шалфеем , колокольчиком и кукурузой .
Женственность
В случае пре-женственности ( протерогинии ) рубец готов к зачатию за некоторое время до опорожнения пыльников. В это время рыльце может опыляться только пыльцой других цветов, что увеличивает вероятность перекрестного опыления. Женственность проявляется, например, в подорожнике .
Несовместимость
Перекрестному опылению также способствует физиологически обусловленная несовместимость между рыльцем и пыльцой или стилевой тканью и пыльцой одного и того же человека: самонесовместимость у растений .
О продолжительности сазаньей тусовки
Если на первой же рыбалке уловистые лунки найдены, то мы на них ловим примерно раз в неделю, не чаще. Замечено, что с каждым следующим выездом наши уловы становятся всё меньше и меньше. В принципе, понятно, почему. Сазан укладывается в лёжку на зиму. Но как долго он это делает? Из года в год мы караулили сазана по первому льду, и статистика показала, что его предзимняя тусовка происходит примерно до Нового года. Лишь однажды нам удалось его поймать в начале января.
Но в тот год и лед встал поздно. Интересно, что последующие рыбалки закончились ничем. Видимо, сазан реагирует именно на температуру воды, а не на наличие льда. Еще мы заметили, чем быстрее приходит зима, тем дольше сазан ловится. То есть, если лед встал в середине ноября или в декабре — всё равно сазан берёт только до Нового года. В принципе, нам этого достаточно.

Очень часто первый лед в Волго-Ахтубинской пойме — это прозрачное стекло, по которому ходишь и смотришь, как рыба разбегается. Ловить хорошего окуня почти бессмысленно. А вот на тех глубинах, где тусуется сазан — пожалуйста, лови! Тут тебе и осторожные поклевки, тут тебе и аккуратное вываживание крупной рыбы, тут тебе и извечный вопрос — пролезет или не пролезет? И еще одно интересное наблюдение – присутствие в прилове карасей. В самом начале караси часто преобладают в улове.
При этом они ловятся не только со дна, но и выше, причем при обыкновенной классической проводке. Но уже через неделю-полторы они куда-то исчезают — и остается только сазан. Так что, если удалось вычислить лунку, где по первому льду ловятся караси – нужно обязательно через пару недель поискать в этих же лунках и сазана. Получается, вырисовываются некоторые правила — и теперь можно систематизировать полученные данные для получения более ясной картины происходящего подо льдом, так что ли?
Способы привлечения опылителей
Выше отмечено, что четыре растения из пяти используют для опыления животных – от млекопитающих до насекомых. Разумеется, при этом они выработали соответствующий механизм для их приманивания.
Самый распространенный – наличие сладкого нектара. Много миллионов лет назад, стремясь приманить к себе опылителей, некоторые растения приобрели клетки, способные выделять сладкое вещество. Опыт оказался удачным, и в результате у него появились многие тысячи потомков, вырабатывавших нектар с резким запахом. Такие растения, в первую очередь, ориентируются на животных и насекомых, обладающих тонким обонянием.

Но ведь есть и растения, использующие орнитофилию. А птицы, как известно, почти лишены обоняния. Здесь был использован иной прием – более крупные и яркие цветы, которые почти не пахнут. Они прекрасно приманивают колибри, которые знают, что внутри их ждет сладкий нектар.
Шаг второй. Изучение исходного материала
Следующим важным шагом будет изучение исходного материала: внутривидовой и видовой потенциал, как искусственный (сорта, гибриды, мутанты и т. д.), так и естественный (разновидности, расы, формы).
Изменчивость живых существ является основой органической эволюции. Изменчивость делится на качественную и количественную. Качественные признаки обычно имеют дискретное значение (опушенный/голый, гладкий/морщинистый), чаще всего контролируются одним или немногими генами и наследуются в соответствии с законами Г. Менделя. Количественные признаки являются суммарным результатом действия многих генов, поддаются подсчетам и измерениям и обладают непрерывной изменчивостью. Проявление количественных признаков сильно зависит от условий среды.
Внутривидовая изменчивость подразумевает, что растения, принадлежащие к одному виду, можно делить на несколько типов категорий. По В. Н. Сукачеву это:
- экотипы (климатипы, эдафотипы, ценотипы) — результат групповой изменчивости;
- лузусы — формы, отличающиеся каким-то признаком, но не обладающие особым ареалом и не принадлежащие к определенным экотипам, а могущие встречаться в разных ареалах и условиях среды;
- аберрации — редкие резко уклоняющиеся формы;
- экады, модификации — ненаследственные формы.
Большое значение имеет закон гомологических рядов, сформулированный Н. И. Вавиловым. Разнообразие форм одного вида или рода можно прогнозировать и у родственных ему видов и родов.
Изучение изменчивости исходных популяций позволяет выбрать наиболее эффективный метод селекции.
При наблюдении за изменчивостью растений важно разграничить влияние среды и наследственности на формирование конкретного фенотипа. Гетерогенность популяций наиболее значительна в области оптимума вида и ниже в экстремальных условиях, однако в экстремальных условиях бывает повышена частота отдельных форм. В изменении отдельных групп признаков могут наблюдаться корреляции
Чем лучше будет изучена внутривидовая изменчивость, её механизмы и особенности в каждом конкретном случае, тем более удачная программа селекции может быть составлена.
Виды перекрестного опыления
На сегодняшний день эксперты выделили у растений две разновидности перекрестного опыления – это ксеногамия и гейтоногамия.
Гейтоногамией называют процесс опыления, в котором задействованы тычинки и пестик разных цветков, расположенных на одном и том же растении. То есть, дерево вполне способно самостоятельно опыляться и давать плоды даже при отсутствии сородичей поблизости.
Поэтому специалисты давно ведут споры о том, является ли это перекрестным опылением или же, скорее, имеет место особый способ самоопыления. Дело в том, что обмена генетическим материалом здесь не происходит, а именно он является главной целью перекрестного опыления.

А вот ксеногамия – совсем другое дело. Процесс проходит точно так же, однако в нем должны быть задействованы цветки, расположенные на разных растениях. Для того, чтобы защищаться от случайного опыления цветков на одном и том же растении, некоторые виды даже выработали защитный механизм – разделение полов. При этом в цветках одного растения находятся только пестики, а на другом – исключительно тычинки. Поэтому вероятность самоопыления полностью исключается.
Приспособления по недопущению самоопыления[править | править код]
Первые покрытосеменные растения, по всей видимости, были обоеполы, что способствовало самоопылению. Позднее растения выработали приспособления по его недопущению.
Разделение половправить | править код
Основная статья: Разделение полов у цветковых растений
Разделение полов — существование растений одного вида, но разного пола: у одних растений образуются цветки только с андроцеем, у других растений — только с гинецеем.
Дихогамияправить | править код
Дихога́мия (от др.-греч. δίχα- «отдельно, врозь» и γάμος «брак») — функциональная разнополость, выраженная в неодновременном созревании в одном цветке андроцея и гинецея; проявляется либо в форме протерандрии, либо в форме протерогинии.
- Протера́ндри́я, или протоа́ндри́я, или прота́ндри́я (от др.-греч. πρότερος «первый из двух, более ранний» и ἀνδρεῖος «мужской»), — созревание в цветке андроцея раньше гинецея (более раннее созревание мужских репродуктивных органов по сравнению с женскими у нецветковых растений; термин применяется также к животным-гермафродитам); протерандрия характерна для семейств Астровые, Зонтичные, Колокольчиковые и многих других.
- Протероги́ни́я, или протоги́ни́я (от др.-греч. πρότερος «первый из двух, более ранний» и γυνή «женщина»), — созревание в цветке гинецея раньше андроцея (более раннее созревание женских репродуктивных органов по сравнению с мужскими у нецветковых растений; термин применяется также к животным-гермафродитам); протерогиния характерна для семейств Барбарисовые, Капустные.
Геркогамияправить | править код
Геркогамия, или херкогамия представляет собой пространственное разделение репродуктивных органов, когда при посещении насекомыми автогамия не может иметь место. Как правило, рыльце пестика выставляется из зева цветка на гораздо большее расстояние, чем тычинки и поэтому самоопыление в форме автогамии кажется невозможным. Тем не менее, у многих геркогамных растений самоопыление часто наблюдается в конце цветения, если по каким-то причинам не произошло перекрестное опыление. При этом происходит изгибание столбиков и тычиночных нитей, нередко обоюдное. В результате репродуктивные органы приходят в соприкосновение в пределах цветка или соцветия. Встречается у лилейных, ирисовых, ластовневых, губоцветных, гераниевых и многих других.
Самонесовместимостьправить | править код
Гетеростилия у первоцвета обыкновенного (Primula vulgaris). 1 — тычинки, 2 — столбики с рыльцами на конце. Слева — длинностолбчатый цветок, справа — короткостолбчатый.
Самонесовместимость — приспособление растений по недопущению самоопыления, выражающееся в том, что при самоопылении число семян ничтожно по сравнению с числом семян при перекрёстном опылении.
Различают гомоморфную и гетероморфную самонесовместимость.
- Гомоморфная самонесовместимость (от др.-греч. ὁμός «одинаковый» и μορφή «форма») — самонесовместимость, не сопровождаемая морфологическими различиями в строении цветка у разных особей одного вида. Характерна, например, для мака самосейки (Papaver rhoeas).
-
Гетеростилия (от др.-греч. ἕτερος — «другой» и στῦλος — «столб»), или Разностолбчатость, или Гетероморфная самонесовместимость (от др.-греч. ἕτερος — «другой» и μορφή «форма»), — самонесовместимость, сочетаемая с существованием особей одного вида, цветки которых имеют различную длину столбиков пестиков и тычиночных нитей (у одних растений столбики короче тычинок, у других — тычинки короче столбиков). Суть этого приспособления состоит в том, что насекомое, касаясь пыльников в цветке одного типа, пачкает своё тело пыльцой в тех местах, которые соответствуют рыльцу столбика в цветке другого типа. Гетеростилия может быть диморфной и триморфной.
- Диморфная гетеростилия — у растений одного вида имеются две формы цветков. Характерна, к примеру, для родов Гречиха (Fagopyrum), Медуница (Pulmonaria), Первоцвет (Primula).
- Триморфная гетеростилия — у растений одного вида имеются три формы цветков: коротко-, средне- и длинностолбчатые. Характерна, к примеру, для дербенника иволистного (плакун-травы) (Lythrum salicaria).
Особенности анемофильных растений
Для всех представителей этой группы характерны такие признаки:
- невзрачные или малозаметные цветки (объясняется тем, что они не должны привлекать насекомых);
- маленькие и сухие пыльцевые зерна;
- большая длина нитей, на которых свисают пыльники.
Теперь подробнее. Главная особенность всех ветроопыляемых культур – это непривлекательность цветков, проявляющаяся в отсутствии нектара, запаха и ярких красок. При этом пыльцевые зерна, которые развиваются в больших количествах, имеют чрезвычайно малые размеры: вес одной пылинки в среднем составляет 0,000001 мг. Приведем небольшое сравнение: пылинка тыквы – растения, опыляемого пчелами – весит в тысячу раз больше, т. е. порядка 0,001 мг. Одно только соцветие конского каштана способно сформировать 42 миллиона зерен, в то время как соцветие ржи – в десять раз меньше (4 миллиона 200 тысяч). К особенностям пыльцы анемофильных растений можно отнести еще и то, что она, будучи полностью лишенной клеящих веществ, зачастую еще и имеет гладкую поверхность.

Конский каштан
Генеративный орган высших растений
Цветок – это видоизмененный побег, где формируются споры и гаметы. Высшие растения (покрытосеменные) имеют сложно устроенные цветки со множеством приспособлений к различным типам опыления. Разнообразный по деталям цветок совмещает в себе процессы как полового, так и бесполого размножения. Главными компонентами цветка являются его репродуктивные части – мужской андроцей (тычинки) и женский гинецей (пестик с завязью, столбик и рыльца). Цветки могут быть обоеполые (есть и пестик, и тычинки) и однополые (есть либо пестик, либо тычинки). Другие составные части цветка очень разнообразны и выполняют специфические функции.